The gene Dual-specificity tyrosine phosphorylation–regulated kinase 1A (DYRK1A) is a highly conserved gene located in the so-called Down Syndrome critical region (DSCR), the part of chromosome 21 thought to be responsible for the majority of phenotypic features in Down syndrome (DS). DYRK1A encodes a dual specificity kinase that phosphorylates diverse regulatory proteins at serine/threonine residues and auto-phosphorylates itself on tyrosine residues to self-activate1,2. DYRK1A regulates many downstream proteins and transcription factors via phosphorylation, it has a wide spectrum of substrates and is involved in many cellular pathways3. DYRK1A substrates are involved in essential cellular functions such as the cell cycle control, cell signalling, apoptosis, chromatin regulation and transcription, RNA splicing, synaptic plasticity, endocytosis and trafficking. This substrate diversity suggests DYRK1A has pleiotropic roles. DYRK1A is a dosage-sensitive enzyme, therefore, both low and high expression exert significant effects3. Consequently, DYRK1A is involved in many pathological disorders including Down syndrome, autism, microcephaly, Alzheimer disease, Parkinson’s disease, Huntington disease and tumorigenesis.
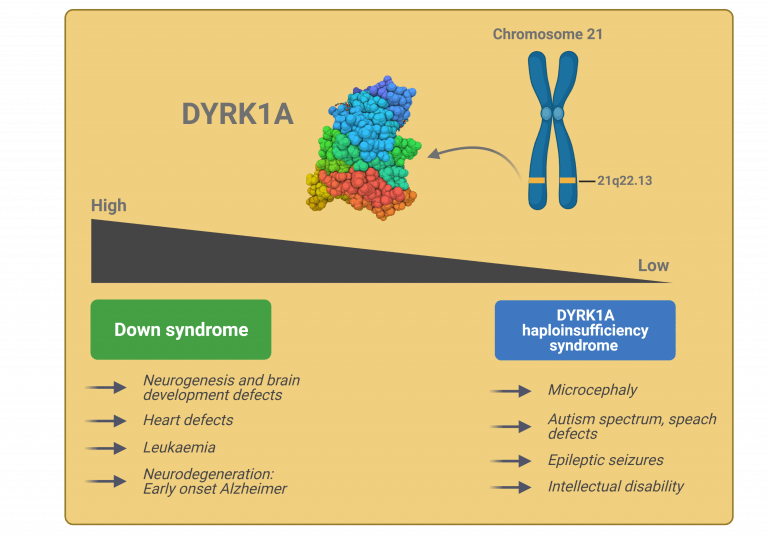
DYRK1A in Down syndrome
DYRK1A is associated with DS characteristics like mental retardation and motor defects.
In 1995, Tejedor and colleagues described a mutant that presented a small brain phenotype due to impaired neural stem cell proliferation. They called the gene minibrain (mnb), and showed that these gene encodes a cell type-specific serine-threonine protein kinase4. A year later, while trying to isolate genes responsible for some features of DS, the team of Nobuyoshi Shimizu identified DYRK1A as the human homolog of mnb5.
Both DYRK1A mRNA and protein expression are increased in DS brain and in the brain of different mouse models of DS6. Similarly, DYRK1A has been found overexpressed in lymphoblastoid cells from patients with DS. Expression levels are on average 1.5 fold higher compared to controls3,7, indicating that the protein is overexpressed in a gene dosage-dependent manner in DS patients. Importantly, studies in mouse models where DYRK1A is triplicated either by overexpressing DYRK1A alone or as part of a genomic segment of chromosome 21 encompassing DYRK1A, have shown that DYRK1A plays a role in the structural, morphological and functional changes that occur in the DS brain, and the associated cognitive and motor deficits8. These models have also shown that overdosage is sufficient to impair cognition and defects are similar to those observed in individuals with DS7.
Evidence suggests that DYRK1A plays a role in the proliferation and differentiation of trisomy 21 neural stem cells9. Knock down of DYRK1A restores the cell cycle (G1) delay found in fibroblast derived from individuals with DS10, and RNAi downregulation improves neuronal differentiation of DS derived iPSCs repectively11.
The involvement of DYRK1A in neural development and function, is related to the regulatory activity that it exerts via phosphorylation of numerous substrates, some of which are described below.
Funded Projects:
Ongoing:
- Roper R. (2020): Correction of Dyrk1a-related Down syndrome skeletal phenotypes during developmentally sensitive periods
- Janel N. (2020): Deciphering the role of DYRK1A and GSK3beta at the crossroads of insulin secretion and sensitivity and cognitive dysfunction in Down syndrome
- Brault V. (2020): Impact of Dyrk1a gene dosage on striatal development and function
- De la Luna S. (2019): Organization of the DYRK1A interactome through docking domains: searching for novel targeting approaches
- Malinge S. (2018): Targeting DYRK1A: a key player in Down syndrome Leukaemogenesis
- Duchon A. (2016): Traitement in utero d’un modèle murin de la Trisomie 21 avec un inhibiteur de Dyrk1a.
Previously funded:
- Pedemonte N. (2017): A drug repositioning approach for the discovery of DYRK1A regulators as a therapy for Down syndrome
- Brault V. (2017): Impact de l’altération de la fonction de Dyrk1a dans les neurones GABAergiques pour l’étude des syndromes de Down et MRD7
- Janel N. (2017): Détermination de biomarqueurs impliqués dans la maladie d’Alzheimer et associés à DYRK1A dans la trisomie 21
- Barallobre M.J. (2017): Identifying new mechanisms by which DYRK1A regulates the expansion of the cerebral cortex
- Malinge S. (2016): Développement de modèles d’études génétiques et à visée thérapeutique pour mesurer l’impact de DYRK1A dans les leucémies associées au syndrome de
- Rueda N. (2016): Effects of the normalization of Dyrk1a copy number on the cerebellar phenotypes and neurodegenerative profile of a mouse model of Down syndrome
- Schenck A. (2014): Spermidine supplementation to improve defects in Drosophila models of DYRK1A and other Intellectual Disability disorders – a pilot study
- Delabar J.M. (2014): Le dosage de DYRK1A agit-il sur les voies impliquées dans la maladie d’Alzheimer dans la trisomie 21?
- Brault V. (2014): Implication du gène Cbs dans le vieillissement cérébral accéléré observé dans le syndrome de Down et interactions avec les gènes Dyrk1A et App
- Herault Y. (2013): Impact de l’altération de la fonction de Dyrk1a au cours dans les compartiments glutamatergiques et gabaergiques dans des modèles souris de la Trisomie 21
- Polak M. (2013): Dyrk1A overexpression and Hypothyroidism in Down Syndrome
- Arbonés de Rafael M.L. (2012): Deciphering how triplication of DYRK1A impacts in early brain neurogenesis
- Dierssen M. (2012): The role of Dyrk1A in Down syndrome obesity: mechanism and therapeutic approaches
- Polak M. (2012): Dyrk1A overexpression and Hypothyroidism in Down Syndrome
- Tejedor F.J. (2012): Involvement of SEPT4 phosphorylation by MNB/DYRK1A in neurite formation and dendritogenesis/synaptogenesis.Implications for Trisomy 21 and pharmacotherapeutics
- Delabar J.M. (2012): Corrélation entre le niveau sérique de Dyrk1a et des marqueurs biochimiques ou cognitifs de la trisomie 21
- Herault Y. (2011): Evaluation de la capacité de nouveaux inhibiteurs de Dyrk1a à améliorer les performances cognitives de modèles souris de la Trisomie 21
- De la Torre Fornell R. (2011): Normalization of dyrk1A function as an approach to improve cognitive performance in SD subjects: Epigallocatechin gallate as therapeutic tool
- London J. (2010): Rôles de la surexpression des gènes APP et Dyrk1a dans les troubles du sommeil et du vieillissement: aspects physiologiques et biochimiques
- Delabar J.M. (2010): Nouveaux inhibiteurs de Dyrk1a
- Polak M. (2010): Dyrk1a (dual-specificity tyrosine-(Y):-phosphorylation regulated kinase 1A):regulation of Shh pathway is linked to Hypothyroidism in Down Syndrome
- Roubertoux P. (2010): Contribution du gène Dscr3 (région D21S17-ETS2): et de ses interactions avec Dyrk1A aux phénotypes cognitifs et cérébraux de la trisomie 21
- De la Torre Fornell R. (2010): EGCG a DYRK1A inhibitor as therapeutic tool for reversing cognitive deficits in Down syndrome individuals
- Manzoni O. (2010): Pharmacothérapie synaptique dans un modèle murin de trisomie 21 les souris Dyrk1A
- Janel N. (2009): Détermination de marqueurs sériques du niveau d’activité de dyrk1a
- Simonneau M. (2009): Anomalies dendritiques induites par le surdosage de dyrk1A: un mecanisme epigénétique impliquant l’interaction de dyrk1A avec une histone déméthylase
- Rodrigues-Lima F. (2009): Etude structurale et enzymatique d’inhibiteurs de Dyrk1A une kinase potentiellement impliquées dans le syndrome de Down
- Tejedor F. J. (2008): Involvement of the MNB/DYRK1A kinase-SEPT4 interaction in Trisomy 21 associated Neuropathologies
- Popowycz F. (2008): Développement d’inhibiteurs de kinase DYRK1A nouvelle approche thérapeutique du syndrome de Down et autres maladies neurodégénératives
- Delabar J.M. (2008): Ciblage thérapeutique de dyrk1a
- Barallobre M.J. (2007): Role of Dyrk1A and DSCR1 during the development of cortical dendrites. Implications in Down Syndrome.
- Yu E. (2007): Effects of human chromosome 21 syntenic regions and Dyrk1a on Trisomy 21-related phenotypes in mice
- Hwang Y.W. (2007): Regulation of the assembly of endocytic complexes by Dyrk1A
- Meijer L. (2007): Inhibiteurs Pharmacologiques de DYRK1A une Protéine Kinase Impliquée dans la Maladie d’Alzheimer et la Trisomie 21
- Fillat Fonts C. (2006): Functional and molecular consequences of inhibiting Dyrk1A gene expression in the hippocampus of Down syndrome mouse models by AAVshDyrk1A delivery.
- Delabar J.M. (2006): Controle pharmacologique de l’action de dyrk1a gène candidat pour la trisomie 21
- Chabert C. (2004): Analyse de nouveaux modèles murins de surexpression des gènes DYRK1A et C21ORF5
- Arbones M.L. (2004): Analyse du role de Dyrk1A un gène candidat pour le retard mental dans la Trisomie 21 dans la neurogénèse
- Bestel A.M. (2003): Etude in vitro & in vivo de la séquence d’expression génique Dyrk1A-Rim impliquée dans la transmission synaptique et modifiée dans la trisomie 21
- Fillat Fons C. (2003): Conséquences phénotypiques de l’utilisation d’ARN interférent ciblé sur Dyrk1a dans des modèles murins de trisomie 21. Implications pour la thérapie.
- Delabar J.M. (2003): Neurogénèse dans des modèles murins surexprimant le gène Dyrk1A
- Chabert C. (2002): Modèles murins de surexpression des gènes DYRK1A et C21ORF5
- Delabar J.M. (2000): Modélisation de la trisomie 21: trois gènes candidats C21ORF5 SIM2 DYRK1A
- Estivill X. (2000): Implication de Dyrk1a dans les défauts d’apprentissage associés a la trisomie 21 et leur corrélations neuropharmacologiques
More information about DYRK1A...
DYRK1A targets and functions
DYRK1A has a nuclear localisation signal and a nuclear speckle targeting signal, but it has been detected in the nucleus, the soma and dendrites of neurons. Consequently, DYRK1A substrates comprise a plethora of nuclear and cytosolic proteins, including transcription factors (CREB, NFAT, Notch, STAT3, FKHR, Gli1), transcriptional regulators (Pol II, Histone H3), splicing factors (cyclin L2, SF2, SF3), cell cycle regulators (p53, cyclin D1, P27KIP1, LIN52), a translation factor (eIF2Be), synaptic proteins (dynamin I, amphiphysin I, synaptojanin I), and others (presenilin, parkin, beta-tubulin, glycogen synthase, caspase-9). Some of the cellular processes in which DYRK1A is involved are3,7,9,12,13 :
Gene expression
DYRK1A plays a role in the regulation of gene expression by targeting a number of chromatin regulators and transcription factors, including:
RNA polymerase II (RNAPII): by phosphorylating the CTD (carboxy-terminal domain) of Pol II at residues necessary for transcriptional elongation, DYRK1A regulates transcription.
Histone H3: by phosphorylating Histone H3, DYRK1A sustains open chromatin structure, activates transcription, prevents heterochromatin protein 1 HP1 from binding and suppressing transcription and participates in the activation of a subset of genes, including those encoding cytokines.
Repressor element-1 binding transcription factor/neuron-restrictive silencer factor (REST/NRSF respectively): important for neural lineage specification, synapse formation and function, seems to be negatively regulated by dosage imbalance of DYRK1A, which promotes its degradation. Thereby, affecting neuronal differentiation.
Nuclear factor of activated T cell cytoplasmic (NFATc): NFATs are transcription factors involved in immune response, tumour progression and development of the cardiac, skeletal muscle and nervous systems. DYRK1A phosphorylates NFATs, regulating their activity by retaining them in the cytoplasm.
Notch: DYRKA can interact with Notch intracellular domain (NICD) in the nucleus, phosphorylate it and preventing the expression of the transcription factor Hes1. Thereby suppressing Notch signalling.
Cell cycle control
DYRK1A is a negative regulator of the cell cycle. It promotes exit of the cell cycle and switch to a quiescent state or differentiation. DYRK1A substrates with a role in cell cycle, include:
p53: involved in cell proliferation by allowing entry into S phase. When phosphorylated by DYRK1A, there is and upregulation of p53 targets, including p21CIP1 which impairs G1/G0-to-S phase transition and hence, inhibiting cell proliferation.
Cyclin D1: involved in cell proliferation by allowing entry into S phase. DYRK1A phosphorylation leads to its nuclear export and degradation by the proteasome, inhibiting cell proliferation and inducing premature differentiation.
P27KIP1: cyclin dependent kinase inhibitor in neural precursors. p27KIP1 upregulation by DYRK1A, inhibits cyclin-dependent kinase complexes that controls G1/S transition, promoting cell cycle exit and neuronal differentiation.
LIN52: component of the DREAM complex that represses cell cycle-dependent genes during quiescence and coordinates periodic gene expression with peaks during the G1/S and G2/M phases. DYRK1A phosphorylation of LIN52 is required for the assembly of the DREAM complex, promoting quiescence and inhibition of cell proliferation.
Cytoskeletal dynamics
DYRK1A can regulate the cytoskeletal architecture by phosphorylation of both tubulin subunits and microtubule-associated proteins. Moreover, it has been shown that APP and tau phosphorylation are mediated by DYRK1A.
Microtubule-associated protein tau: Phosphorylation of tau by DYRK1A reduces the ability of tau to promote microtubule assembly and stability,
Microtubule-associated protein 1b (Map1b): phosphorylation of Map1b by DYRK1a, primes this protein for subsequent phosphorylation by Gsk3-beta which is involved in the regulation of microtubules stability in growing cortical neuronal axons.
β-tubulin: Phosphorylation of β-tubulin by DYRK1A inhibits tubulin polymerization and microtubules dynamics, which is critical for dendritic patterning and neuronal function.
Ciliogenesis
Recent studies demonstrated that DYRK1A is expressed in ciliated tissues, localizes to ciliary axonemes and basal bodies, and is required for ciliogenesis14. Furthermore, a putative role of DYRK1A during mitosis was suggested given its localisation to mitotic spindles. These two processes require dynamic microtubules remodelling, process in which DYRK1A also plays a role (see above).
Neurogenesis and neuronal maturation
In 1995, Tejedor and colleagues published the first evidence that DYRK1A was involved in brain growth. In their paper, they described a Drosophila mutant that presented a small brain phenotype due to impaired neural stem cell proliferation. They called the gene minibrain (mnb)4, later identified as the Drosophila homolog of DYRK1A. After that, many studies have shown that the function of DYRK1A in brain growth is conserved across evolution. In humans, truncating mutations in the DYRK1A gene, cause microcephaly15–17 and autism18, and both mouse and zebrafish models with loss-of-function mutations of DYRK1A, also present microcephaly. Compelling data from work in several models have shown that DYRK1A regulates neural proliferation and differentiation19 via the above mentioned DYRK1A targets and functions. Thus, DYRK1A plays a role in neurogenesis. Importantly, alterations in neurogenesis have an impact in size and organization of the brain, its circuitry and in neuronal plasticity, all of which are essential for brain function and cognition.
DYRK1A implication in neurodegeneration
Finally, there are increasing evidences that DYRK1A is involved with neurodegeneration, and participates in the pathogenesis of several neurodegenerative diseases such as Alzheimer, Parkinson’s and Huntington disease. This is due to the fact that it phosphorylates several proteins related to these diseases, including tau, alpha-synuclein, septin-4, presenilin or amyloid beta precursor protein (APP). Furthermore, it accumulates in amyloid plaques.
Thus, given the involvement of DYRK1A in neural development and brain functions, and neurodegenerative processes that are altered in DS (e.g. neurofibrillary degeneration, neuronal loss and Alzheimer disease-like phenotype), this kinase has received considerable attention20 and is one of the current candidates for targeted interventions aimed at improving brain development and cognition in individuals with DS. Importantly, development of drugs targeting DYRK1A, will also help in the treatment of neurodegenerative diseases.
References
- Walte, A. et al. Mechanism of dual specificity kinase activity of DYRK1A. FEBS J. 280, 4495–4511 (2013).
- Kentrup, H. et al. Dyrk, a dual specificity protein kinase with unique structural features whose activity is dependent on tyrosine residues between subdomains VII and VIII. J. Biol. Chem. 271, 3488–3495 (1996).
- Laham, A. J., Saber-Ayad, M. & El-Awady, R. DYRK1A: a down syndrome-related dual protein kinase with a versatile role in tumorigenesis. Cell. Mol. Life Sci. CMLS (2020) doi:10.1007/s00018-020-03626-4.
- Tejedor, F. et al. minibrain: a new protein kinase family involved in postembryonic neurogenesis in Drosophila. Neuron 14, 287–301 (1995).
- Shindoh, N. et al. Cloning of a human homolog of the Drosophila minibrain/rat Dyrk gene from ‘the Down syndrome critical region’ of chromosome 21. Biochem. Biophys. Res. Commun. 225, 92–99 (1996).
- Dowjat, W. K. et al. Trisomy-driven overexpression of DYRK1A kinase in the brain of subjects with Down syndrome. Neurosci. Lett. 413, 77–81 (2007).
- Arbones, M. L., Thomazeau, A., Nakano-Kobayashi, A., Hagiwara, M. & Delabar, J. M. DYRK1A and cognition: A lifelong relationship. Pharmacol. Ther. 194, 199–221 (2019).
- Herault, Y. et al. Rodent models in Down syndrome research: impact and future opportunities. Dis. Model. Mech. 10, 1165–1186 (2017).
- Stagni, F., Giacomini, A., Emili, M., Guidi, S. & Bartesaghi, R. Neurogenesis impairment: An early developmental defect in Down syndrome. Free Radic. Biol. Med. 114, 15–32 (2018).
- Chen, J.-Y., Lin, J.-R., Tsai, F.-C. & Meyer, T. Dosage of Dyrk1a Shifts Cells within a p21-Cyclin D1 Signaling Map to Control the Decision to Enter the Cell Cycle. Mol. Cell 52, 87–100 (2013).
- Hibaoui, Y. et al. Modelling and rescuing neurodevelopmental defect of Down syndrome using induced pluripotent stem cells from monozygotic twins discordant for trisomy 21. EMBO Mol. Med. 6, 259–277 (2014).
- Feki, A. & Hibaoui, Y. DYRK1A Protein, A Promising Therapeutic Target to Improve Cognitive Deficits in Down Syndrome. Brain Sci. 8, (2018).
- Abbassi, R., Johns, T. G., Kassiou, M. & Munoz, L. DYRK1A in neurodegeneration and cancer: Molecular basis and clinical implications. Pharmacol. Ther. 151, 87–98 (2015).
- Willsey, H. R. et al. The neurodevelopmental disorder risk gene DYRK1A is required for ciliogenesis and control of brain size in Xenopus embryos. Dev. Camb. Engl. 147, (2020).
- Courcet, J.-B. et al. The DYRK1A gene is a cause of syndromic intellectual disability with severe microcephaly and epilepsy. J. Med. Genet. 49, 731–736 (2012).
- Møller, R. S. et al. Truncation of the Down syndrome candidate gene DYRK1A in two unrelated patients with microcephaly. Am. J. Hum. Genet. 82, 1165–1170 (2008).
- Ji, J. et al. DYRK1A haploinsufficiency causes a new recognizable syndrome with microcephaly, intellectual disability, speech impairment, and distinct facies. Eur. J. Hum. Genet. 23, 1473–1481 (2015).
- van Bon, B. W. M. et al. Disruptive de novo mutations of DYRK1A lead to a syndromic form of autism and ID. Mol. Psychiatry 21, 126–132 (2016).
- Tejedor, F. J. & Hämmerle, B. MNB/DYRK1A as a multiple regulator of neuronal development. FEBS J. 278, 223–235 (2011).
- Duchon, A. & Herault, Y. DYRK1A, a Dosage-Sensitive Gene Involved in Neurodevelopmental Disorders, Is a Target for Drug Development in Down Syndrome. Front. Behav. Neurosci. 10, 104 (2016).